
Lipid


| Tipovi masti u hrani |
|---|
|
| Vidi još |
Lipidi (grč. λίπος — mast) jesu jedinjenja različitog sastava po pravilu nerastvorna u vodi, a rastvorna u organskim rastvaračima.[4] Biološki su veoma značajna jedinjenja. Oni su osnovna komponenta bioloških membrana i utiču na njihovu propustljivost, učestvuju u predaji nervnih impulsa, stvaraju kontakte među ćelijama, čine energetske rezerve, štite organizam od mehaničkih povreda i formiraju termoizolacioni sloj.[5]
Zbog specifične hemijske strukture, lipidi se sastoje od nepolarnog i polarnog dela. Oni su često uprošćeno nazivani mastima, ali taj naziv nije ispravan jer su lipidi grupa koja ne obuhvata samo jedinjenja masnih kiselina i glicerola, već i drugih supstanci. Dakle, masti su u stvari samo jedna od nekoliko grupa lipida.[6]
Pregled[uredi | uredi izvor]
Lipidi obuhvataju veliku grupu raznorodnih jedinjenja sa zajedničkom odlikom nerastvorljivosti u vodi, i rastvorljivosti u organskim rastvaračima: benzolu, hloroformu, acetonu, vrelom alkoholu. Lipidi u biljnim i životinjskim organizmima učestvtvuju kao gradivne i rezervne materije. U metabolizmu lipida značajnu ulogu ima aktivirana sirćetna kiselina (ili acetil-koenzim-acetil-KoA). Prema strukturnoj organizaciji, masti se dele na:
- proste i
- složene masti.[7]
Prosti lipidi su estri trihidroksilnog alkohola (glicerola) i masnih kiselina (oleinske, pamitinske i stearinske). Čvrste masti (loj) čine jedinjenja glicerola sa zasićenom palmitinskom (C16H36O2) ili stearinskom (C18H36O2) kiselinom. Za razliku od njih, u sastav tečnih masti ili ulja ulazi oleinska (C17H34O2). Proste masti sa vodom čine nestabilne emulzije.
Zagrevanjem masti u prisustvu alkalija (NaOH ili KOH) dolazi do saponifikacije, čime se dobijaju jedinjenja sapuna (koja su rastvorna u vodi). Procesom saponifikacije masti se razgrađuju na glicerol i soli masnih kiselina. Uvođenjem vodonika u nezasićene masne kiseline (procesom hidrogenizacije), one prelaze u zasićene sa izmenjenim mirisom i ukusom. Zahvaljujući primeni ovog postupka, u industriji se razvio postupak pravljenja margarina i biljnog masla.
Lipidi, kao rezervna organsku materija, su karakteristični po velikoj energetskoj vrednost, a počinju se trošiti samo u nedostatku glukoze. Višak ugljenih hidrata u organizmu se pretvara u masti i skladišti u različitim organima (kod životinja, npr. najčešće se odlažu potkožno). Takav slučaj je posebno značajan kod ptica selica i životinja sa sezonskim zimskim snom.
Voskovi, za razliku od glicetida, umesto glicerola sadrže dugolančane jednovalentne alkohole. Pored alkohola, sadrže i više masne kiseline. Zastupljeni su u biljkama i životinjama. Nisu rastvorljivi u vodi, već u organskim rastvaračima. Među poznatijim supstancama ove grupe su: pčelinji vosak, spermacet i lanolin.
U složene lipide ubrajaju se fosfolipidi, glikolipidi i lipoproteini. Izgrađeni su od glicerola, masnih kiselina i fosforne kiseline.[7]
Od poznatijih fosfolipida, najčešći su lecitin, kefalin, plazmalogen i lizofosfolipid. Fosfolipidi čine temeljnu komponentu žive ćelijske membrane i ostalih ćelijskih uldopina. Odvijanje normalnih životnih procesa u organizmu, vezano je za specifične funkcije složenih lipida. Tako je za kontrahovanje glatke muskulature neophodan medijator acetilholin. On takođe ima poseban značaj u podraživanju parasimpatičkih nerava, te snižavanje krvnog pritiska. Fosfolipidi takođe učestvuju u permeabilnosti ćelijskih (živih) membrana, posebno pri prenosu jona Na+ i K+ u srčanom mišiću (miokardu), dok od nekih drugih zavisi permeabilnost membrane mitohondrija. Lizofosfatidi izazivaju hemolizu (razgradnju) eritrocita. U zmijskom otrovu nalazi se fosfolipaza A, zbog čega nakon ujeda dolazi do hemolize eritrocita.
Glikolipidi obuhvataju cerebrozide, gangliozide, sulfatide itd. Cerebrozidi su prvi put izolovani iz mozga (cerebrum), po čemu su i dobili ime. Gangliozidi su izolovani iz ganglijskih ćelija mozga (sive mase). Sulfatidi su jedinjenja složenih lipida izolovana iz jetre i bubrega, a relativno visokim sadržajem ovih glikolipida obogaćen je i mozak. Sva ova jedinjenja u organizmima životinja imaju vrlo specifične i značajne funkcije.[7]
Klasifikacija i osobine[uredi | uredi izvor]
Prosti lipidi su supstance čiji se molekuli sastoje samo od ostataka masnih kiselina i alkohola (najčešće glicerola). Ovde spadaju masti i ulja (trigliceridi) i voskovi.[8][9]
Složeni lipidi uključuju derivate fosforne kiseline (fosfolipidi) i lipide koji sadrže ostatke ugljenih hidrata (glikolipidi). Ovde spadaju i steroidi.[10]
Ako se životinjska ili biljna tkiva tretiraju organskim rastvaračima, deo će se rastvoriti. Komponente rastvora se nazivaju lipidi. Lipidna frakcija sadrži supstance različitih tipova.
Masne kiseline spadaju u alifatične karboksilne kiseline. Iz lipida je izolovano više od 70 raznih masnih kiselina. Dele se na zasićene i nezasićene. Prirodne masne kiseline sadrže paran broj C atoma, a niz nije razgranat, već je normalan. Ispod su nabrojane neke od najvažnijih masnih kiselina (masne se zovu zato što se nalaze u mastima):
| Zasićene masne kiseline | |
| Trivijalni naziv | Strukturna formula |
| buterna | CH3(CH2)2COOH |
| kapronska | CH3(CH2)4COOH |
| kaprilna | CH3(CH2)6COOH |
| kaprinska | CH3(CH2)8COOH |
| laurinska | CH3(CH2)10COOH |
| miristinska | CH3(CH2)12COOH |
| palmitinska | CH3(CH2)14COOH |
| stearinska | CH3(CH2)16COOH |
| arahidinska | CH3(CH2)18COOH |
| begenska | CH3(CH2)20COOH |
| lignocerinska | CH3(CH2)22COOH |
| Nezasićene masne kiseline | |
| Trivijalni naziv | Strukturna formula |
| krotonska | CH3 - CH = CH - COOH |
| palmatooleinska | CH3 - (CH2)5 - CH = CH - (CH2)7 - COOH |
| oleinska | CH3 - (CH2)7 - CH = CH - (CH2)7 - COOH |
| elaidinska | CH3 - (CH2)7 - CH = CH - (CH2)11 - COOH |
| nervonska | CH3 - (CH2)7 - CH = CH - (CH2)13 - COOH |
| linolna | CH3 - (CH2)4 - CH = CH - CH2 - CH = CH - (CH2)7 - COOH |
| linoleinska | CH3 - CH2 - CH = CH - CH2 - CH = CH - CH2 - CH = CH - (CH2)7 - COOH |
| arahidonska | CH3 - (CH2)4 - CH = CH - CH2 - CH = CH - CH2 - CH = CH - CH2 - CH = CH - (CH2)3 - COOH |
| klupadonska | CH3 - CH2 - CH = CH - (CH2)2 - CH = CH - CH2 - CH = CH - (CH2)2 - CH = CH - (CH2)2 - CH = CH - (CH2)2 - COOH |
Najvažnije prirodne masne kiseline sadrže 16 ili 18 C atoma. Nezasićene masne kiseline obično sadrže dvostruke veze između C9 i C10. Ostale dvostruke veze nisu konjugovane i nalaze se prema CH3- kraju niza. Imaju cis- konfiguraciju, a veoma retko trans konfiguraciju. Masne kiseline sa većim brojem C atoma su praktično nerastvorne u vodi. Njihove Na i K- soli se zovu sapuni. One obrazuju micele u vodi kod kojih he negativan COO- kraj okrenut ka vodi, a nepolaran C- niz je okrenut ka unutrašnjosti micele. U celini micele su naelektrisane negativno i u rastvoru ostaju suspendovane jer se međusobno odbijaju.
Gliceridi[uredi | uredi izvor]
Neutralne masti (gliceridi) (acilgliceridi) su estri glicerola (glicerina) i masnih kiselina. U odnosu na broj estarskih veza u molekulu razlikuju se monogliceridi, digliceridi i trigliceridi (odnosno, monoacilglicerini, diacilglicerini i triacilglicerini).
Masti se u organizmima nalaze kao protoplazmatične masti (u određenoj količini) i kao rezervne masti (u neodređenoj količini). Osnovnu masu masti čine trigliceridi. Ako su sva tri kiselinska ostatka ista, prosti trigliceridi (na primer, tripalmitin), a ako su ostaci od od različitih kiselina, mešoviti trigliceridi. Kod davanja naziva, cifre ukazuju na kome se C atomu nalazi odgovarajući kiselinski ostatak (na primer. 1-oleo-2-pal-mitostearin).
Ukoliko su kiselinski ostaci duži, utoliko raste i tačka topljenja. Ako su kiseline nezasićene, tačka topljenja sa brojem dvostrukih veza opada. Životinjske masti sadrže znatnu količinu zasićenih masnih kiselina (naročito stearinsku i palmitinsku) pa su na sobnoj temperaturi čvrste. Ako se u molekulu nalazi dosta nezasićenih kiselina to su onda ulja, koja su na sobnoj temperaturi tečna (maslinovo ulje, sojino, suncokretovo, ulje uljane repice). Mast kod ljudi ima tačku topljenja 15 °C i sadrži 70% oleinske kiseline.
Gliceridi reaguju kao estri, a posebno je važna reakcija hidrolize, pri čemu se izdvaja glicerin i masne kiseline. Hidroliza se može vršiti enzimski ili dejstvom kiselina ili baza. Bazna hidroliza sa natrijum hidroksidom (NaOH) ili sa kalijum hidroksidom (KOH) se koristi za proizvodnju sapuna (reakcija saponifikacije, sapo-sapun).
Masti sadrže razne primese koje se uklanjaju, a zaostaje tzv. neutralna mast. Ona se karakteriše:
- jodnim brojem (broj grama joda koga vezuje 100 g masti pod određenim uslovima i koji karakteriše stepen nezasićenosti. Jodni broj svinjske masti je 46-66, a maslinovog ulja oko 85).
- kiselinski broj (broj mg KOH potreban za neutralizaciju 1 g masti i koji ukazuje na količinu slobodnih masnih kiselina u masti).
- saponifikacioni broj (broj mg KOH potreban za neutralizaciju svih kiselina – i slobodnih i vezanih u trigliceride – u 1 g masti).
Voskovi[uredi | uredi izvor]
Voskovi su estri viših masnih kiselina i viših alkohola sa 20 do 70 C atoma u molekulu. Oni se nalaze u sastavu masti koja pokriva kožu, vunu, perje. Kod biljaka su 80% svih lipida voskovi koji stvaraju zaštitni sloj na površini biljke i štite je od isparavanja vode.
Prirodni vosak (pčelinji, lanolin) pored pomenutih estara sadrži i malu količinu slobodnih masnih kiselina, nekih alkohola i ugljovodonika, sa 20-35 C atoma u molekulu. Opšta formula voskova je:
Fosfolipidi[uredi | uredi izvor]
Fosfolipidi ili fosfatidi se dele na glicerofosfolipide i sfingolipide.
- Glicerofosfolipide su derivati fosfatidne kiseline i sadrže glicerol, masne kiseline, fosfatidnu kiselinu i azotna jedinjenja. Njihova opšta formula je:

gde su R i R' ostaci viših masnih kiselina, a R" je ostatak nekog azotnog jedinjenja i fosforne kiseline.
Jedan deo molekula (R i R') je jako hidrofoban, a drugi deo je hidrofilan (negativno naelektrisanje u fosfornoj kiselini i pozitivno naelektrisanje azotnog jedinjenja – R"). Od svih lipida glicerofosfolipidi su najpolarniji, a u vodenoj sredini se nalazi u obliku micela. Ima nekoliko grupa glicerofosfolipida:
- Fosfatidilholini, lecitini sadrže glicerol, više masne kiseline, fosfornu kiselinu i holin:

- Fosfatidiletanolamini, kefalini umesto holina sadrže etanolamin:

Gornje dve grupe su glavna lipidna komponenta membrane ćelija.
- Fosfatidilserini kao azotna jedinjenja sadrže ostatak serina:

Od masnih kiselina koje ulaze u sastav glicerofosfolipida najčešće su stearinska, palmitinska, oleinska i linolenska. Pri njihovoj hidrolizi (otrov kobre) izdvaja se ostatak nezasićene masne kiseline, a preostalo jedinjenje ima jako hemolitičko dejstvo.
Sfingolipidi[uredi | uredi izvor]
- Sfingolipidi ne sadrže glicerol već alkohol sfingozin:
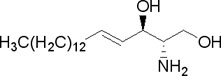

Najčešće su sfingomijelini koji se nalaze u membranama ćelija i to posebno nervnih. Hidrolizom daju jedan molekul masne kiseline, jedan molekul nezasićenog dvovalentnog alkohola sfingozina, jedan molekul azotnog jedinjenja (najčešće holina) i jedan molekul fosforne kiseline. Hemijska formula sfingomijelina data je ispod:

Polarni deo je naelektrisan i pozitivno (holin) i negativno (ostatak fosforne kiseline), a ostali deo molekula je nepolaran.
Glikolipidi[uredi | uredi izvor]
Glikolipidi su složeni lipidi koji sadrže ostatak ugljenih hidrata (najčešće D-galaktoze). Bitni su za rad bioloških membrana. Nalaze se u mozgu i krvi. Dele se ne dve grupe:
- Cerebrozidi sadrže heksozu (često D-galaktozu) vezanu etarskom vezom sa sfingozinom, a sadrže i neku masnu kiselinu sa 24 C atoma (lignocerinska, cerebronska):

Mnogo cerebrozida se nalazi u membranama nervnih ćelija.
Gangliozidi[uredi | uredi izvor]
Gangliozidi hidrolizom daju višu masnu kiselinu, sfingozin, D-glukozu, D-galaktozu kao i derivate aminošećera: N-acetilglukozamin i N-acetilneuraminsku kiselinu. Nalaze se pretežno u sivoj kori mozga i membranama nervih ćelija.
Svi gore pomenuti lipidi spadaju u saponifikujuće lipide jer pri njihovoj baznoj hidrolizi nastaju sapuni. Postoje i lipidi koji se ne hidrolizuju uz oslobađanje masnih kiselina. U tu grupu spadaju steroidi.
Biološke funkcije[uredi | uredi izvor]
Membrane[uredi | uredi izvor]

Eukariotske ćelije su kompartmentalizovane u organele obavijene membranama, koje vrše različite biološke funkcije. Glicerofosfolipidi su glavna strukturna komponenta bioloških membrana, kao što su ćelijska membrana i intraćelijske membrane organela; u životinjskim ćelijama membrane fizički odvajaju intraćelijske komponente od ekstracelularnog okruženja. Glicerofosfolipidi su amfipatički molekuli (sadrže hidrofobne i hidrofilne regione) koji imaju glicerolno jezgro vezano sa dve masne kiseline estarskim vezama, i za jednu „čeonu“ grupu putem fosfatne estarske veze. Dok su glicerofosfolipidi glavne komponente bioloških membrana, druge negliceridne lipidne komponente kao što su sfingomijelin i steroli (uglavnom holesterol u životinjskim ćelijskim membranama) su isto tako prisutni u biološkim membranama.[11] Kod biljki i algi galaktozildiacilgliceroli[12] i sulfohinovozildiacilglicerol,[13] kojima nedostaje fosfatna grupa, su važne komponente membrana hloroplasta i srodnih organela i najzastupljeniji su lipidi u fotosintetičkim tkivima, uključujući ona kod viših biljki, algi i pojedinih bakterija.
Najveća lipidna komponenta biljne tilakoidne membrane je monogalaktozil diglicerid (MGDG) koji obično ne formira dvosloj, dok su fosfolipidi prisutni u manjoj meri. Uprkos toj jedinstvenoj lipidnoj kompoziciji, za hloroplastne tilakoidne membrane je putem magnetne rezonance i elektronske mikroskopije pokazano da sadrže dinamičku lipidni dvoslojnu matricu.[14]
Biološka membrana je forma lamelarne faze lipidnog dvosloja. Formiranje lipidnog dvosloja je energetski preferentan proces kad su gore opisani glicerofosfolipidi u vodenom okruženju.[15] To je poznato kao hidrofobni efekat. U vodenoj sredini, polarne glave lipida se poravnavaju ka polarnom, vodenom okruženju, dok hidrofobni repovi minimizuju svoj kontakt sa vodom i grupišu se zajedno, formirajući vezikule; u zavisnosti od koncentracije lipida, ta biofizička interakcija može da dovede do formiranja micela, lipozoma, ili lipidnih dvosloja. Drugi oblici agregacije su isto tako primećeni i formiraju deo polimorfizma amfifilnog (lipidnog) ponašanja. Fazno ponašanje je oblast izučavanja u okviru biofizike i predmet je tekućih akademskih istraživanja.[16][17] Micele i dvoslojevi se formiraju u polarnom medijumu procesu poznatom kao hidrofobni efekat.[18] Pri rastvaranju lipofilne ili amfifilne supstance u polarnoj sredini, polarni molekuli (i.e., voda u vodenim rastvorima) postaju u većoj meri uređeni oko rastvorene lipofilne supstance, pošto polarni molekuli ne mogu da formiraju vodonične veze sa lipofilnim oblastima amfifila. Stoga u vodenom okruženju, molekuli vode formiraju uređene "klatratne" kaveze oko rastvorenih lipofilnih molekula.[19]
Organizovanje lipida u protoćelijske membrane je ključni korak u modelima abiogeneze, porekla života.[20]
Čuvanje energije[uredi | uredi izvor]
Trigliceridi, uskladišteni u adipoznom tkivu, su glavna forma energetskog skladišta kod životinja i biljki. Adipocit, ili masna ćelija, je dizajniran za kontinuiranu sintezu i razlaganje triglicerida kod životinja, pri čemu se razlaganje uglavnom aktivira enzimima senzitivnim na hormone, lipazama.[21] Kompletna oksidacija masnih kiselina pruža visok kalorijski sadržaj, oko 9 kcal/g, u poređenju sa 4 kcal/g za razlaganje ugljenih hidrata i proteina. Migratorne ptice mogu da lete preko velikih rastojanja bez konzumiranja hrane koristeći energiju sačuvanu u obliku triglicerida kao gorivo za svoj let.[22]
Signalizacija[uredi | uredi izvor]
Zadnjih godina je postalo jasno da je lipidna signalizacija vitalni deo ćelijske signalizacije.[23][24] Lipidna signalizacija se može odvijati putem aktivacije G protein spregnutih ili nuklearnih receptora, i članovi nekoliko različitih kategorija lipida su identifikovani kao signalni molekuli i ćelijski glasnici.[25] Među njima su sfingozin-1-fosfat, sfingolipid izveden iz keramida koji je potentan glasnički molekul koji učestvuje u regulaciji kalcijumske mobilizacije,[26] u ćelijskom rastu, i apoptozi;[27] diacilglicerol (DAG) i fosfatidilinozitol fosfati (PIP), koji učestvuju u kalcijumom posredovanoj aktivaciji proteinske kinaze C;[28] prostaglandini, koji su jedan od tipova iz masnih kiselina izvedenih eikosanoida koji učestvuju u inflamaciji i imunosti;[29] steroidni hormoni kao što su estrogen, testosteron i kortizol, koji modulišu domaćinove funkcije reprodukcije, metabolizma i krvnog pritiska; i oksisteroli kao što je 25-hidroksi-holesterol koji je agonist jetrenog X receptora.[30] Poznato je da fosfatidilserinski lipidi učestvuju u signalizaciji fagocitoze apoptotičkih ćelija i/ili delova ćelija. Oni ostvaruju taj cilj tako što su izloženi na ekstracelularnom licu ćelijske membrane nakon inaktivacije flipaza koje ih ekskluzivno postavljaju na citozolnu stranu i aktivacije skramblaza, koje izokreću orijentaciju fosfolipida. Nakon toga druge ćelije prepoznaju fosfatidilserine i fagocitoziraju ćelije ili ćelijske fragmente koji ih izlažu.[31]
Druge funkcije[uredi | uredi izvor]
Vitamini rastvorni u mastima (A, D, E i K) – koji su na izoprenu bazirana jedinjenja – su esencijalni nutrijenti koji se čuvaju u jetri i masnim tkivima, sa raznovrsnim opsegom funkcija. Acil-karnitini učestvuju u transportu i metabolizmu masnih kiselina „u i iz“ mitohondrija, gde oni podležu beta oksidaciji.[32] Poliprenoli i njihovi fosforilisani derivati takođe imaju važnu transportne uloge, u tom slučaju u transportu oligosaharida kroz membrane. Poliprenolni fosfatni šećeri i poliprenolni difosfatni šećeri uzimaju učešća u reakcijama glikozilacije izvan citoplazme, u biosintezi ekstracelularnih polisaharida (na primer, peptidoglikanskoj polimerizaciji kod bakterija), i u N-glikozilaciji eukariotskih proteina.[33][34] Kardiolipini su potklasa glicerofosfolipida koji sadrže četiri acilna lanca i tri glicerolne grupe, koja je posebno izobilna u unutrašnjoj mitohondrijskoj membrani.[35][36] Smatra se da oni aktiviraju enzime koji učestvuju u oksidativnoj fosforilaciji.[37] Lipidi isto tako formiraju osnovu steroidnih hormona.[38]
Metabolizam[uredi | uredi izvor]
Glavni prehrambeni lipidi kod ljudi i životinja su biljni trigliceridi, steroli, i membranski fosfolipidi. Procesom lipidnog metabolizma se sintetišu i degradiraju uskladišteni lipidi i proizvode se strukturni i funkcionalni lipidi karakteristični za pojedinačna tkiva.
Biosinteza[uredi | uredi izvor]
Kod životinja, kad postoji višak ponude prehrambenih ugljenih hidrata, suvišni ugljeni hidrati se konvertuju u trigliceride. Taj proces obuhvata sintezu masnih kiselina iz acetil-KoA i esterifikaciju masnih kiselina radi produkcije triglicerida. Ovaj proces se naziva lipogeneza.[39] Masne kiseline se formiraju sintezom masnih kiselina, kojom se polimerizuju i zatim redukuju acetil-KoA jedinice. Acilni lanci masnih kiselina se produžavaju ciklusom reakcija koje dodaju acetil grupu, redukuju je do alkohola, dehidratišu alkohol do alkenske grupe i zatim se ponovo vrši redukcija do alkanske grupe. Enzimi masno kiselinske biosinteze se dele u dve grupe, kod životinja i gljiva sve reakcije sinteze masnih kiselina se izvode posredstvom jednog multifunkcionalnog proteina,[40] dok u biljnim plastidima i bakterijama zasebni enzimi vrše svaki korak tog procesa.[41][42] Masne kiseline se mogu naknadno konvertovati do triglicerida koji se pakuju u lipoproteinima i izlučuju iz jetre.
Sinnteza nezasićenih masnih kiseline obuhvata reakciju desaturacije, pri čemu se dvostruka veza uvodi u masni acilni lanac. Na primer, kod ljudi, desaturacija stearinske kiseline posredstvom enzima stearoil-KoA desaturaza-1 proizvodi oleinsku kiselinu. Dvostruko nezasićena masna kiselina linoleinska kiselina, kao i trostruko nezasićena α-linoleinska kiselina se ne mogu sintetisati u tkivima sisara, i stoga su esencijalne masne kiseline i moraju se unositi kao hrana.[43]
Trigliceridna sinteza se odvija u endoplazmatičnom retikulumu putem metaboličkih puteva u kojima se acilne grupe u masnim acil-KoA molekulima prenose na hidroksine grupe glicerol-3-fosfata i diacilglicerola.[44]
Terpeni i izoprenoidi, uključujući karotenoide, se formiraju putem pravljenja i modifikacije izoprenskih jedinica iz reaktivnih prekursora izopentenil pirofosfata i dimetilalil pirofosfata.[45] Ti prekursori mogu da budu napravljeni na različite načine. Kod životinja i arheja, mevalonatni putem se formiraju ta jedinjenja iz acetil-KoA,[46] dok se u biljkama i bakterijama u nemevalonatnom putu koriste pyruvat i gliceraldehid 3-fosfat kao supstrati.[45][47] Jedna važna reakcija koja koristi te aktivirane izoprenske donore je steroidna biosinteza. Ovde se izoprenske jedinice spajaju da bi se formirao skvalen i zatim se savijaju da bi nastala grupa prstenova koji sačinjavaju lanosterol.[48] Lanosterol se može dalje konvertovati u druge steroide kao što je holesterol i ergosterol.[48][49]
Degradacija[uredi | uredi izvor]
Beta oksidacija je metabolički proces kojim se masne kiseline razlažu u mitohondrijama i/ili u peroksizomima da bi se formirao acetil-KoA. Najvećim delom, masne kiseline se oksiduju putem mehanizma koji je sličan sa, mada ne i identičan sa, reverznim procesom sinteze masnih kiselina. Drugim rečima, dvougljenični fragmenti se uklanjaju sekvencijalno sa karboksilnog kraja kiseline nakon koraka dehidrogenacije, hidracije, i oksidacije uz formiranje beta-keto kiseline, koje se odvaja tiolizom. Acetil-KoA se zatim ultimatno konvertuje u ATP, CO2, i H2O koristeći ciklus limunske kiseline i lanac elektronskog transporta. Ciklus limunske kiseline može da počne od acetil-KoA kad se masnoća razlaže radi energije, ako je prisutno malo ili je odsutna glukoza. Energetski prinos od kompletne oksidacije masne kiseline palmitata je 106 ATP.[50] Za degradaciju nezasićenih i neparnih masnih kiselina neophodni su dodatni enzimatski procesi.
Reference[uredi | uredi izvor]
- ^ Maitland 1998, str. 139
- ^ Stryer et al.. str. 328.
- ^ Stryer et al.. str. 330.
- ^ Clayden, Jonathan; Greeves, Nick; Warren, Stuart; Wothers, Peter (2001). Organic Chemistry (I izd.). Oxford University Press. ISBN 978-0-19-850346-0.
- ^ David L. Nelson; Michael M. Cox (2005). Principles of Biochemistry (IV izd.). New York: W. H. Freeman. ISBN 0-7167-4339-6.
- ^ Šerban 2010.
- ^ a b v Međedović, S.; Maslić, E.; Hadžiselimović, R. (2002). Biologija 2. Sarajevo: Svjetlost. ISBN 978-9958-10-222-6.
- ^ Koraćević, Darinka; Bjelaković, Gordana; Đorđević, Vidosava. Biohemija. Savremena administracija. ISBN 978-86-387-0622-8.
- ^ autora, Grupa (2006). Ugrešić, Nenad, ur. Farmakoterapijski vodič 3 (PDF). Beograd. ISSN 1451-4680. Arhivirano iz originala (PDF) 15. 07. 2011. g. Pristupljeno 23. 05. 2010.
- ^ Keith Parker; Laurence Brunton; Goodman, Louis Sanford; Lazo, John S.; Gilman, Alfred (2006). Goodman & Gilman's The Pharmacological Basis of Therapeutics (11. izd.). New York: McGraw-Hill. ISBN 0071422803.
- ^ Stryer et al.. str. 329–331.
- ^ Heinz E. "Plant glycolipids: structure, isolation and analysis". str. 211–332 in Advances in Lipid Methodology, Vol. 3. W.W. Christie (ed.). Oily Press. Dundee. 1996. ISBN 978-0-9514171-6-4.
- ^ Hölzl, G.; Dörmann, P. (2007). „Structure and function of glycoglycerolipids in plants and bacteria”. Progress in Lipid Research. 46 (5): 225—243. PMID 17599463. doi:10.1016/j.plipres.2007.05.001.
- ^ Yashroy, RC. (1990). „Magnetic resonance studies of dynamic organisation of lipids in chloroplast membranes”. Journal of Biosciences. 15 (4): 281—288. doi:10.1007/BF02702669.
- ^ Stryer et al.. str. 333–334.
- ^ van Meer G, Voelker DR, Feigenson GW (2008). „Membrane lipids: where they are and how they behave”. Nature Reviews Molecular Cell Biology. 9 (2): 112—124. PMC 2642958
 . PMID 18216768. doi:10.1038/nrm2330.
. PMID 18216768. doi:10.1038/nrm2330.
- ^ Feigenson, GW. (2006). „Phase behavior of lipid mixtures”. Nature Chemical Biology. 2 (11): 560—563. PMC 2685072 . PMID 17051225. doi:10.1038/nchembio1106-560.
- ^ Wiggins, PM. (1990). „Role of water in some biological processes”. Microbiological Reviews. 54 (4): 432—449. PMC 372788 . PMID 2087221.
- ^ Raschke, T. M.; Levitt, M. (2005). „Nonpolar solutes enhance water structure within hydration shells while reducing interactions between them”. Proceedings of the National Academy of Sciences of the United States of America. 102 (19): 6777—6782. PMC 1100774 . PMID 15867152. doi:10.1073/pnas.0500225102.
- ^ Segré, D.; Ben-Eli D; Deamer, D.; Lancet D (2001). „The Lipid World” (PDF). Origins of Life and Evolution of Biospheres. 31 (1–2): 119—145. PMID 11296516. doi:10.1023/A:1006746807104. Arhivirano iz originala (PDF) 11. 09. 2008. g. Pristupljeno 13. 02. 2017.
- ^ Brasaemle, DL. (2007). „Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis”. Journal of Lipid Research. 48 (12): 2547—2559. PMID 17878492. doi:10.1194/jlr.R700014-JLR200.
- ^ Stryer et al.. str. 619.
- ^ „Lipid”. Current Opinion in Plant Biology. 7 (3): 329—236. 2004. PMID 15134755. doi:10.1016/j.pbi.2004.03.012.
|first1=zahteva|last1=u Authors list (pomoć) - ^ Dinasarapu, A. R.; Saunders, B.; Ozerlat I; Azam K, Subramaniam S (2011). „Signaling gateway molecule pages—a data model perspective”. Bioinformatics. 27 (12): 1736—1738. PMC 3106186 . PMID 21505029. doi:10.1093/bioinformatics/btr190.
- ^ Eyster, KM. (2007). „The membrane and lipids as integral participants in signal transduction”. Advances in Physiology Education. 31 (1): 5—16. PMID 17327576. doi:10.1152/advan.00088.2006.
- ^ Hinkovska-Galcheva V; VanWay, S. M.; Shanley, T. P.; Kunkel, R. G. (2008). „The role of sphingosine-1-phosphate and ceramide-1-phosphate in calcium homeostasis”. Current Opinion in Investigational Drugs. 9 (11): 1191—1205. PMID 18951299.
- ^ Saddoughi, S. A.; Song, P.; Ogretmen, B. (2008). „Roles of bioactive sphingolipids in cancer biology and therapeutics”. Subcellular Biochemistry. Subcellular Biochemistry. 49: 413—440. ISBN 978-1-4020-8830-8. PMC 2636716 . PMID 18751921. doi:10.1007/978-1-4020-8831-5_16.
- ^ Klein, C.; Malviya, A. N. (2008). „Mechanism of nuclear calcium signaling by inositol 1,4,5-trisphosphate produced in the nucleus, nuclear located protein kinase C and cyclic AMP-dependent protein kinase”. Frontiers in Bioscience. 13 (13): 1206—1226. PMID 17981624. doi:10.2741/2756.
- ^ Boyce, JA. (2008). „Eicosanoids in asthma, allergic inflammation, and host defense”. Current Molecular Medicine. 8 (5): 335—349. PMID 18691060. doi:10.2174/156652408785160989.
- ^ „Liver”. Cardiovascular Therapy. 26 (4): 297—316. 2008. PMID 19035881. doi:10.1111/j.1755-5922.2008.00062.x.
|first1=zahteva|last1=u Authors list (pomoć) - ^ Biermann, M.; Maueröder C; Brauner JM,Chaurio R, Janko C, Herrmann M, Muñoz LE (2013). „Surface code—biophysical signals for apoptotic cell clearance”. Physical Biology. 10 (6): 065007. PMID 24305041. doi:10.1088/1478-3975/10/6/065007.
- ^ Indiveri, C; Tonazzi, A; Palmieri, F (1991). „Characterization of the unidirectional transport of carnitine catalyzed by the reconstituted carnitine carrier from rat liver mitochondria”. Biochimica et biophysica acta. 1069 (1): 110—6. PMID 1932043.
- ^ Parodi, A. J.; Leloir, L. F. (1979). „The role of lipid intermediates in the glycosylation of proteins in the eucaryotic cell”. Biochimica et Biophysica Acta. 559 (1): 1—37. PMID 375981. doi:10.1016/0304-4157(79)90006-6.
- ^ Helenius, A.; Aebi M (2001). „Intracellular functions of N-linked glycans”. Science. 291 (5512): 2364—2369. PMID 11269317. doi:10.1126/science.291.5512.2364.
- ^ Nowicki, M.; Frentzen M (2005). „Cardiolipin synthase of Arabidopsis thaliana”. FEBS Letters. 579 (10): 2161—2165. PMID 15811335. doi:10.1016/j.febslet.2005.03.007.
- ^ Gohil, V. M.; Greenberg, M. L. (2009). „Mitochondrial membrane biogenesis: phospholipids and proteins go hand in hand”. Journal of Cell Biology. 184 (4): 469—472. PMC 2654137 . PMID 19237595. doi:10.1083/jcb.200901127.
- ^ Hoch, FL. (1992). „Cardiolipins and biomembrane function”. Biochimica et Biophysica Acta. 1113 (1): 71—133. PMID 10206472. doi:10.1016/0304-4157(92)90035-9.
- ^ Steroids Arhivirano na sajtu Wayback Machine (23. oktobar 2011). Elmhurst.edu. Pristupljeno 2013-10-10.
- ^ Stryer et al.. str. 634.
- ^ Chirala, S.; Wakil S (2004). „Structure and function of animal fatty acid synthase”. Lipids. 39 (11): 1045—1053. PMID 15726818. doi:10.1007/s11745-004-1329-9.
- ^ White, S. W.; Zheng, J.; Zhang, Y. M.; Rock, C. O. (2005). „The structural biology of type II fatty acid biosynthesis”. Annual Review of Biochemistry. 74: 791—831. PMID 15952903. doi:10.1146/annurev.biochem.74.082803.133524.
- ^ Ohlrogge, J.; Jaworski J (1997). „Regulation of fatty acid synthesis”. Annual Review of Plant Physiology and Plant Molecular Biology. 48: 109—136. PMID 15012259. doi:10.1146/annurev.arplant.48.1.109.
- ^ Stryer, str. 643
- ^ Stryer et al.. str. 733–739.
- ^ a b Kuzuyama, T.; Seto H (2003). „Diversity of the biosynthesis of the isoprene units”. Natural Product Reports. 20 (2): 171—183. PMID 12735695. doi:10.1039/b109860h.
- ^ Grochowski, L.; Xu H; White, R. (2006). „Methanocaldococcus jannaschii uses a modified mevalonate pathway for biosynthesis of isopentenyl diphosphate”. Journal of Bacteriology. 188 (9): 3192—3198. PMC 1447442 . PMID 16621811. doi:10.1128/JB.188.9.3192-3198.2006.
- ^ „The”. Annual Review of Plant Physiology and Plant Molecular Biology. 50: 47—65. 1999. PMID 15012203. doi:10.1146/annurev.arplant.50.1.47.
|first1=zahteva|last1=u Authors list (pomoć) - ^ a b „Sterol”. Annual Review of Biochemistry. 50: 585—621. 1981. PMID 7023367. doi:10.1146/annurev.bi.50.070181.003101.
|first1=zahteva|last1=u Authors list (pomoć) - ^ Lees, N.; Skaggs B; Kirsch, D.; Bard M (1995). „Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—a review”. Lipids. 30 (3): 221—226. PMID 7791529. doi:10.1007/BF02537824.
- ^ Stryer et al.. str. 625–626.
Literatura[uredi | uredi izvor]
- Maitland, Jr Jones (1998). Organic Chemistry. W W Norton & Co Inc (Np). str. 139. ISBN 978-0-393-97378-5.
- Međedović, S.; Maslić, E.; Hadžiselimović, R. (2002). Biologija 2. Svjetlost, Sarajevo. ISBN 978-9958-10-222-6.
- autora, Grupa (2006). Ugrešić, Nenad, ur. Farmakoterapijski vodič 3 (PDF). Beograd. ISSN 1451-4680. Arhivirano iz originala (PDF) 15. 07. 2011. g. Pristupljeno 23. 05. 2010.
- Koraćević, Darinka; Bjelaković, Gordana; Đorđević, Vidosava. Biohemija. Savremena administracija. ISBN 978-86-387-0622-8.
- Šerban, Nada M. (2010). Biologija 1. Beograd: ZUNS. str. 11. ISBN 978-86-17-17585-4.
- Bhagavan, NV (2002). Medical Biochemistry. San Diego: Harcourt/Academic Press. ISBN 978-0-12-095440-7.
- Devlin, T. M. (1997). Textbook of Biochemistry: With Clinical Correlations (4th izd.). Chichester: John Wiley & Sons. ISBN 978-0-471-17053-2.
- Stryer, L.; Berg, J. M.; Tymoczko, J. L. (2007). Biochemistry (6th izd.). San Francisco: W.H. Freeman. ISBN 978-0-7167-8724-2.
- van Holde KE, Mathews CK (1996). Biochemistry (2nd izd.). Menlo Park, California: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-3931-4.
- Georg Löffler, Petro E. Petrides (2003). Biochemie und Pathobiochemie. Berlin: Springer. ISBN 978-3-540-42295-2.
- Florian Horn, Isabelle Moc, Nadine Schneider: Biochemie des Menschen. Thieme. . Stuttgart. 2005. ISBN 978-3-13-130883-2.
- Charles E. Mortimer, Ulrich Müller: Chemie. Thieme. . Stuttgart. 2003. ISBN 978-3-13-484308-8.
- F.D. Gunstone, J.L. Harwood, F.B. Padley: The Lipid Handbook. Chapman and Hall, London New - York. 1986. ISBN 978-0-412-24480-3.
- O.W. Thiele: Lipide, Isoprenoide mit Steroiden. G. . Stuttgart: Thieme Verlag. 1979. ISBN 978-3-13-576301-9.
- Robert C. Murphy: Mass Spectrometry of Lipids, Handbook of Lipid Research Vol. 7, Plenum Press. New York and London. 1993. ISBN 978-0-306-44361-9.
- Julian N. Kanfer and Sen-itiroh Hakomori, Sphingolipid Biochemistry, vol. 3 of Handbook of Lipid Research (1983)
- Dennis E. Vance and Jean E. Vance (eds.), Biochemistry of Lipids and Membranes (1985).
- Donald M. Small, The Physical Chemistry of Lipids, vol. 4 of Handbook of Lipid Research (1986).
- Robert B. Gennis, Biomembranes: Molecular Structure and Function (1989)
- Charles R. Scriver, Arthur L. Beaudet, William S. Sly, and David Valle, The Metabolic and Molecular Bases of Inherited Disease (1995).
- Gunstone, F. D. Fatty acids and lipid chemistry. — London: Blackie Academic and Professional, 1996. 252 pp.
- Robert M. Bell, John H. Exton, and Stephen M. Prescott (eds.), Lipid Second Messengers, vol. 8 of Handbook of Lipid Research (1996).
- Christopher K. Mathews, K.E. van Holde, and Kevin G. Ahern, Biochemistry, 3rd ed. (2000).
- Alberts, B., et al. «Essential Cell Biology, » Garland Science. (2nd изд.). 2004. ISBN 978-0-8153-3480-4..
- Solomon, Eldra P., et. al. (2005) «Biology, 7th Edition.» Thomson, Brooks/Cole.
- «Advanced Biology — Principles and Applications.» C.J. Clegg and D.G. Mackean. ISBN 978-0-7195-7670-6..
- Fahy E. et al. A comprehensive classification system for lipids // J. Lipid. Res. 2005. V. 46, № 5. P. 839—861.
Spoljašnje veze[uredi | uredi izvor]
- ApolloLipids - Provides dyslipidemia and cardiovascular disease prevention and treatment information as well as continuing medical education programs.
- Lipids, Membranes and Vesicle Trafficking - The Virtual Library of Biochemistry and Cell Biology
- The Lipid library - provides information on the chemistry, analysis and biochemistry of lipids
- LIPID MAPS: LIPID Metabolites and Pathways Strategy
- IUPAC glossary entry for the lipid class of molecules what is IUPAC?
- "A comprehensive classification system for lipids" Arhivirano na sajtu Wayback Machine (24. avgust 2010)
- CCMDWeb, a lipid health educational resource on global risk reduction for dyslipidemia and cardiovascular disease as well as online CME programs.
- European Federation for the Science and Technology of Lipids