
Transpiracija
1 - Voda se pasivno transportuje u korene i zatim u ksilem.
2 - Dejstvom sila kohezije i adhezije se formira stub molekula vode u ksilemu.
3 - Voda prelazi iz ksilema u mezofilne ćelije, isparava sa njihove površine i napušta biljku putem difuzije kroz stome.


Transpiracija je proces odavanja vode (u vidu vodene pare) sa površine biljaka, naročito sa površine listova.[1] Postoje tri osnovna tipa transpiracije: kutikularna - odigrava se sa površine cele biljke, molekuli vode prolaze kroz kutikulu, lenticelarna - odigrava se kroz otvore na stablu koji se nazivaju lenticele, i stomaterna - kontrolisano odavanje vode kroz stome. Objedinjuje se sa procesom evaporacije u naziv evapotranspiracija, kojim se označava važna faza u kruženju vode u prirodi.[2]
Voda je neophodna biljkama, ali se samo mala količina vode koju uzimaju koreni koristi za rast i metabolizam. Preostalih 97–99,5% se gubi transpiracijom i gutacijom.[3] Površine listova su prošarane porama koje se nazivaju stomate (jednina „stoma”). Kod većine biljaka one su brojnije na donjoj strani lišća. Stomate su oivičene zaštitnim ćelijama i pomoćnim ćelijama stomata (koje su zajedno poznate kao stomatalni kompleks). One otvaraju i zatvaraju pore.[4] Transpiracija se dešava kroz otvore stomata i može se smatrati neophodnim „troškom“ povezanom sa otvaranjem stomata kako bi se omogućila difuzija gasa ugljen-dioksida iz vazduha za fotosintezu. Transpiracija takođe hladi biljke, menja osmotski pritisak ćelija i omogućava masovni protok mineralnih hranljivih materija i vode od korena do izdanaka. Dva glavna faktora utiču na brzinu protoka vode od tla do korena su: hidraulička provodljivost zemljišta i veličina gradijenta pritiska kroz zemljište. Oba ova faktora utiču na brzinu protoka vode koja se kreće od korena do stomatalnih pora u listovima preko ksilema.[2]
Masovni protok tečne vode od korena do listova delimično je vođen kapilarnim delovanjem, ali prvenstveno zbog razlika u potencijalu vode. Ako je potencijal vode u ambijentalnom vazduhu manji od potencijala vode u vazdušnom prostoru lista stomatalnih pora, vodena para će putovati niz gradijent i kretati se iz vazdušnog prostora lista u atmosferu. Ovo kretanje smanjuje potencijal vode u vazdušnom prostoru lista i izaziva isparavanje tečne vode sa zidova ćelija mezofila. Ovo isparavanje povećava napetost vodenih meniskusa u ćelijskim zidovima i smanjuje njihov radijus, a samim tim i napon koja se vrši nad vodom u ćelijama. Zbog kohezivnih svojstava vode, napon se kreće kroz ćelije lista do ksilema lista i stabljike gde se stvara trenutni negativan pritisak dok voda povlači ksilem iz korena.[5] Kako se isparavanje dešava na površini lista, svojstva adhezije i kohezije rade u tandemu kako bi izvukli molekule vode iz korena, kroz tkivo ksilema i iz biljke kroz stomate.[6] Kod viših biljaka i drveća sila gravitacije se može prevazići jedino smanjenjem hidrostatskog (vodenog) pritiska u gornjim delovima biljaka usled difuzije vode iz stomata u atmosferu. Voda se apsorbuje u korenu osmozom, a svi rastvoreni mineralni hranljivi sastojci putuju sa njom kroz ksilem.
Teorija kohezije i napetosti objašnjava kako lišće povlači vodu kroz ksilem. Molekuli vode se drže zajedno ili ispoljavaju koheziju. Kako molekul vode isparava sa površine lista, on povlači susedni molekul vode, stvarajući neprekidan tok vode kroz biljku.[7]
Kapilarno delovanje[uredi | uredi izvor]
Kapilarno delovanje je proces tečenja tečnosti u uskim prostorima bez pomoći, ili čak u suprotnosti sa spoljnim silama poput gravitacije. Efekat se može videti u uvlačenju tečnosti između dlačica četkice, u tankoj cevi, u poroznim materijalima kao što su papir i gips, u nekim neporoznim materijalima kao što su pesak i tečna ugljenična vlakna, ili u biološka ćelija. To nastaje zbog međumolekulskih sila između tečnosti i okolnih čvrstih površina. Ako je prečnik cevi dovoljno mali, onda kombinacija površinskog napona (koja je uzrokovana kohezijom unutar tečnosti) i sila adhezije između tečnosti i zida posude deluje da pokreće tečnost.[8]
Regulacija[uredi | uredi izvor]
Biljke regulišu brzinu transpiracije kontrolisanjem veličine stomatalnih otvora. Na brzinu transpiracije takođe utiče potreba za isparavanjem atmosfere koja okružuje list, kao što je provodljivost graničnog sloja, vlažnost, temperatura, vetar i sunčeva svetlost. Zajedno sa nadzemnim faktorima, temperatura zemljišta i vlaga mogu uticati na otvaranje stomata,[9] a time i na brzinu transpiracije. Količina vode koju izgubi biljka takođe zavisi od njene veličine i količine vode koja se apsorbuje u korenu. Transpiracija predstavlja najveći deo gubitka vode od strane biljke lišćem i mladim stabljikama. Transpiracija služi za hlađenje biljaka isparavanjem, jer voda koja isparava odnosi toplotnu energiju zbog svoje velike latentne toplote isparavanja od 2260 kJ po litru.
| Karakteristika | Efekat na transpiraciju |
|---|---|
| Broj listova | Više listova (ili bodlji, ili drugih organa za fotosintezu) znači veću površinu i više stomata za razmenu gasova. To dovodi do većeg gubitka vode. |
| Broj stomata | Više stomata će obezbediti više pora za transpiraciju. |
| Veličina lista | List sa većom površinom će se transpirisati brže od lista sa manjom površinom. |
| Prisustvo biljne kutikule | Voštana kutikula je relativno nepropusna za vodu i vodenu paru i smanjuje isparavanje sa površine biljke osim preko stomata. Reflektujuća kutikula može da smanji sunčevo zagrevanje i porast temperature lista, pomažući da se smanji stopa isparavanja. Sićušne strukture slične dlakama koje se nazivaju trihomi na površini listova takođe mogu inhibirati gubitak vode stvaranjem okruženja visoke vlažnosti na površini listova. Ovo su neki primeri prilagođavanja biljaka za očuvanje vode koji se mogu naći na mnogim kserofitima. |
| Snabdevanje svetlošću | Brzina transpiracije kontroliše se stomatalnim otvorom, a ove male pore se otvaraju posebno za fotosintezu. Iako postoje izuzeci od ovoga (kao što je noćna ili CAM fotosinteza), generalno, snabdevanje svetlom će podstaći otvorene stomate. |
| Temperatura | Temperatura utiče na brzinu na dva načina:
1) Povećana brzina isparavanja usled porasta temperature će ubrzati gubitak vode. |
| Relativna vlažnost | Sušnije okruženje daje strmiji gradijent potencijala vode i tako povećava brzinu transpiracije. |
| Vetar | U mirnom vazduhu, voda izgubljena usled transpiracije može se akumulisati u obliku pare blizu površine lista. Ovo smanjuje stopu gubitka vode, pošto je gradijent potencijala vode od unutrašnjeg ka spoljašnjem delu lista tada nešto manji. Vetar oduva veliki deo ove vodene pare blizu površine lista, čineći potencijalni gradijent strmijim i ubrzavajući difuziju molekula vode u okolni vazduh. Čak i na vetru, međutim, može doći do akumulacije vodene pare u tankom graničnom sloju vazduha koji se sporije kreće pored površine lista. Što je vetar jači, to će ovaj sloj težiti da bude tanji, a gradijent potencijala vode strmiji. |
| Snabdevanje vodom | Nedostatak vode uzrokovan ograničenim snabdevanjem vodom iz tla može dovesti do zatvaranja stomata i smanjenja stope transpiracije. |
-
 Uticaj temperature na brzinu transpiracije biljaka.
Uticaj temperature na brzinu transpiracije biljaka. -
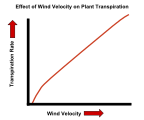 Uticaj brzine vetra na brzinu transpiracije biljaka.
Uticaj brzine vetra na brzinu transpiracije biljaka. -
 Uticaj vlage na brzinu transpiracije biljaka.
Uticaj vlage na brzinu transpiracije biljaka.




Tokom sezone rasta, list transpiriše mnogo puta više vode od sopstvene težine. Jutar kukuruza ispušta oko 3.000–4.000 galona (11.400–15.100 litara) vode svakog dana, a veliki hrast može da ispusti 40.000 galona (151.000 litara) godišnje. Stopa transpiracije je odnos mase transpirirane vode prema masi proizvedene suve materije; koeficijent transpiracije useva ima tendenciju da padne između 200 i 1000 (i.e. biljke useva transpiruju 200 do 1000 kg vode za svaki kg proizvedene suve materije).[10]
Stope transpiracije biljaka mogu se meriti brojnim tehnikama, uključujući potometere, lizimetre, porometre, sisteme fotosinteze i termometričke senzore protoka soka. Merenja izotopa pokazuju da je transpiracija veća komponenta evapotranspiracije.[11] Nedavni dokazi iz jedne globalne studije[12] stabilnih izotopa u vodi pokazuju da se transpirisana voda izotopski razlikuje od podzemnih voda i potoka. Ovo sugeriše da voda u tlu nije tako dobro izmešana kao što se široko pretpostavlja.[13]
Kavitacija[uredi | uredi izvor]
Da bi se održao gradijent pritiska neophodan da bi biljke ostale zdrave, one moraju kontinuirano da unosi vodu svojim korenima. One moraju da budu u stanju da zadovolje potrebe vode izgubljene zbog transpiracije. Ako biljka nije u stanju da unese dovoljno vode da ostane u ravnoteži sa transpiracijom, dolazi do pojave poznate kao kavitacija.[14] Kavitacija se javlja kada biljka ne može da snabde svoj ksilem adekvatnom količinom vode, te umesto da se ispuni vodom, ksilem počinje da se puni vodenom parom. Ove čestice vodene pare se spajaju i formiraju blokade unutar ksilema biljke. Ovo sprečava biljku da transportuje vodu kroz svoj vaskularni sistem.[15] Ne postoji očigledan obrazac gde se kavitacija javlja u ksilemu biljke. Ako se taj problem efikasno ne razreši, kavitacija može dovesti do toga da biljka dostigne svoju trajnu tačku uvenuća i umre. Prema tome, biljka mora da ima metod kojim će ukloniti ovu blokadu kavitacije, ili mora da stvori novu vezu vaskularnog tkiva u celoj biljci.[16] Biljka to čini zatvaranjem svojih stomata preko noći, što zaustavlja tok transpiracije. Ovo onda omogućava korenima da generišu pritisak od preko 0,05 mPa, a to je u stanju da uništi blokadu i ponovo napuni ksilem vodom, ponovo povezujući vaskularni sistem. Ako biljka nije u stanju da stvori dovoljan pritisak da iskoreni blokadu, ona mora sprečiti širenje blokade upotrebom kruške koštice, a zatim stvoriti novi ksilem koji može ponovo da poveže vaskularni sistem biljke.[17]
Reference[uredi | uredi izvor]
- ^ Cummins, Benjamin (2007), Biological Science (3 izd.), Freeman, Scott, str. 215
- ^ a b Taiz, Lincoln (2015). Plant Physiology and Development. Sunderland, MA: Sinauer Associates, Inc. str. 101. ISBN 978-1-60535-255-8.
- ^ Sinha, Rajiv Kumar (2004-01-01). Modern Plant Physiology (na jeziku: engleski). CRC Press. ISBN 978-0-8493-1714-9.
- ^ Benjamin Cummins (2007), Biological Science (3 izd.), Freeman, Scott, str. 215
- ^ Freeman, Scott (2014). Biological Sciences. United States of America: Pearson. str. 765—766. ISBN 978-0-321-74367-1.
- ^ Simon, E.J., Dickey, J.L, & Reece, J.B. (2019). Campbell essential biology. 7th New York: Pearson
- ^ Graham, Linda E. (2006). Plant Biology. Upper Saddle River, NJ 07458: Pearson Education, Inc. str. 200–202. ISBN 978-0-13-146906-8.
- ^ „Capillary Action – Liquid, Water, Force, and Surface – JRank Articles”. Science.jrank.org. Arhivirano iz originala 2013-05-27. g. Pristupljeno 2013-06-18.
- ^ Mellander, Per-Erik; Bishop, Kevin; Lundmark, Tomas (2004-06-28). „The influence of soil temperature on transpiration: a plot scale manipulation in a young Scots pine stand”. Forest Ecology and Management (na jeziku: engleski). 195 (1): 15—28. ISSN 0378-1127. doi:10.1016/j.foreco.2004.02.051.
- ^ Martin, J.; Leonard, W.; Stamp, D. (1976), Principles of Field Crop Production (3rd izd.), New York: Macmillan Publishing Co., ISBN 978-0-02-376720-3
- ^ Jasechko, Scott; Sharp, Zachary D.; Gibson, John J.; Birks, S. Jean; Yi, Yi; Fawcett, Peter J. (3. 4. 2013). „Terrestrial water fluxes dominated by transpiration”. Nature. 496 (7445): 347—50. Bibcode:2013Natur.496..347J. PMID 23552893. S2CID 4371468. doi:10.1038/nature11983.
- ^ Evaristo, Jaivime; Jasechko, Scott; McDonnell, Jeffrey J. (2015-09-03). „Global separation of plant transpiration from groundwater and streamflow”. Nature. 525 (7567): 91—94. Bibcode:2015Natur.525...91E. ISSN 0028-0836. PMID 26333467. S2CID 4467297. doi:10.1038/nature14983.
- ^ Bowen, Gabriel (2015-09-03). „Hydrology: The diversified economics of soil water”. Nature. 525 (7567): 43—44. Bibcode:2015Natur.525...43B. ISSN 0028-0836. PMID 26333464. S2CID 205086035. doi:10.1038/525043a.
- ^ Zhang, Yong-Jiang (decembar 2016). „Reversible Leaf Xylem Collapse: A Potential "Circuit Breaker" against Cavitation”. Plant Physiology. 172 (4): 2261—2274. PMC 5129713
 . PMID 27733514. doi:10.1104/pp.16.01191.
. PMID 27733514. doi:10.1104/pp.16.01191.
- ^ Hochberg, Uri (jun 2017). „Stomatal Closure, Basal Leaf Embolism, and Shedding Protect the Hydraulic Integrity of Grape Stems”. Plant Physiology. 174 (2): 764—775. PMC 5462014 . PMID 28351909. doi:10.1104/pp.16.01816.
- ^ Holbrook, Michele (maj 2001). „In Vivo Observation of Cavitation and Embolism Repair Using Magnetic Resonance Imaging”. Plant Physiology. 126 (1): 27—31. PMC 1540104 . PMID 11351066. doi:10.1104/pp.126.1.27.
- ^ Tiaz, Lincoln (2015). Plant Physiology and Development. Massachusetts: Sinauer Associates, Inc. str. 63. ISBN 978-1605352558.
Spoljašnje veze[uredi | uredi izvor]
- Transpiration by Trees
- USGS The Water Cycle: Evapotranspiration Arhivirano na sajtu Wayback Machine (7. decembar 2013)
| Državne | |
|---|---|
| Ostale | |