
Транспирација
1 - Вода се пасивно транспортује у корене и затим у ксилем.
2 - Дејством сила кохезије и адхезије се формира стуб молекула воде у ксилему.
3 - Вода прелази из ксилема у мезофилне ћелије, испарава са њихове површине и напушта биљку путем дифузије кроз стоме.


Транспирација је процес одавања воде (у виду водене паре) са површине биљака, нарочито са површине листова.[1] Постоје три основна типа транспирације: кутикуларна - одиграва се са површине целе биљке, молекули воде пролазе кроз кутикулу, лентицеларна - одиграва се кроз отворе на стаблу који се називају лентицеле, и стоматерна - контролисано одавање воде кроз стоме. Обједињује се са процесом евапорације у назив евапотранспирација, којим се означава важна фаза у кружењу воде у природи.[2]
Вода је неопходна биљкама, али се само мала количина воде коју узимају корени користи за раст и метаболизам. Преосталих 97–99,5% се губи транспирацијом и гутацијом.[3] Површине листова су прошаране порама које се називају стомате (једнина „стома”). Код већине биљака оне су бројније на доњој страни лишћа. Стомате су оивичене заштитним ћелијама и помоћним ћелијама стомата (које су заједно познате као стоматални комплекс). Оне отварају и затварају поре.[4] Транспирација се дешава кроз отворе стомата и може се сматрати неопходним „трошком“ повезаном са отварањем стомата како би се омогућила дифузија гаса угљен-диоксида из ваздуха за фотосинтезу. Транспирација такође хлади биљке, мења осмотски притисак ћелија и омогућава масовни проток минералних хранљивих материја и воде од корена до изданака. Два главна фактора утичу на брзину протока воде од тла до корена су: хидрауличка проводљивост земљишта и величина градијента притиска кроз земљиште. Оба ова фактора утичу на брзину протока воде која се креће од корена до стоматалних пора у листовима преко ксилема.[2]
Масовни проток течне воде од корена до листова делимично је вођен капиларним деловањем, али првенствено због разлика у потенцијалу воде. Ако је потенцијал воде у амбијенталном ваздуху мањи од потенцијала воде у ваздушном простору листа стоматалних пора, водена пара ће путовати низ градијент и кретати се из ваздушног простора листа у атмосферу. Ово кретање смањује потенцијал воде у ваздушном простору листа и изазива испаравање течне воде са зидова ћелија мезофила. Ово испаравање повећава напетост водених менискуса у ћелијским зидовима и смањује њихов радијус, а самим тим и напон која се врши над водом у ћелијама. Због кохезивних својстава воде, напон се креће кроз ћелије листа до ксилема листа и стабљике где се ствара тренутни негативан притисак док вода повлачи ксилем из корена.[5] Како се испаравање дешава на површини листа, својства адхезије и кохезије раде у тандему како би извукли молекуле воде из корена, кроз ткиво ксилема и из биљке кроз стомате.[6] Код виших биљака и дрвећа сила гравитације се може превазићи једино смањењем хидростатског (воденог) притиска у горњим деловима биљака услед дифузије воде из стомата у атмосферу. Вода се апсорбује у корену осмозом, а сви растворени минерални хранљиви састојци путују са њом кроз ксилем.
Теорија кохезије и напетости објашњава како лишће повлачи воду кроз ксилем. Молекули воде се држе заједно или испољавају кохезију. Како молекул воде испарава са површине листа, он повлачи суседни молекул воде, стварајући непрекидан ток воде кроз биљку.[7]
Капиларно деловање[уреди | уреди извор]
Капиларно деловање је процес течења течности у уским просторима без помоћи, или чак у супротности са спољним силама попут гравитације. Ефекат се може видети у увлачењу течности између длачица четкице, у танкој цеви, у порозним материјалима као што су папир и гипс, у неким непорозним материјалима као што су песак и течна угљенична влакна, или у биолошка ћелија. То настаје због међумолекулских сила између течности и околних чврстих површина. Ако је пречник цеви довољно мали, онда комбинација површинског напона (која је узрокована кохезијом унутар течности) и сила адхезије између течности и зида посуде делује да покреће течност.[8]
Регулација[уреди | уреди извор]
Биљке регулишу брзину транспирације контролисањем величине стоматалних отвора. На брзину транспирације такође утиче потреба за испаравањем атмосфере која окружује лист, као што је проводљивост граничног слоја, влажност, температура, ветар и сунчева светлост. Заједно са надземним факторима, температура земљишта и влага могу утицати на отварање стомата,[9] а тиме и на брзину транспирације. Количина воде коју изгуби биљка такође зависи од њене величине и количине воде која се апсорбује у корену. Транспирација представља највећи део губитка воде од стране биљке лишћем и младим стабљикама. Транспирација служи за хлађење биљака испаравањем, јер вода која испарава односи топлотну енергију због своје велике латентне топлоте испаравања од 2260 kJ по литру.
| Карактеристика | Ефекат на транспирацију |
|---|---|
| Број листова | Више листова (или бодљи, или других органа за фотосинтезу) значи већу површину и више стомата за размену гасова. То доводи до већег губитка воде. |
| Број стомата | Више стомата ће обезбедити више пора за транспирацију. |
| Величина листа | Лист са већом површином ће се транспирисати брже од листа са мањом површином. |
| Присуство биљне кутикуле | Воштана кутикула је релативно непропусна за воду и водену пару и смањује испаравање са површине биљке осим преко стомата. Рефлектујућа кутикула може да смањи сунчево загревање и пораст температуре листа, помажући да се смањи стопа испаравања. Сићушне структуре сличне длакама које се називају трихоми на површини листова такође могу инхибирати губитак воде стварањем окружења високе влажности на површини листова. Ово су неки примери прилагођавања биљака за очување воде који се могу наћи на многим ксерофитима. |
| Снабдевање светлошћу | Брзина транспирације контролише се стоматалним отвором, а ове мале поре се отварају посебно за фотосинтезу. Иако постоје изузеци од овога (као што је ноћна или CAM фотосинтеза), генерално, снабдевање светлом ће подстаћи отворене стомате. |
| Температура | Температура утиче на брзину на два начина:
1) Повећана брзина испаравања услед пораста температуре ће убрзати губитак воде. |
| Релативна влажност | Сушније окружење даје стрмији градијент потенцијала воде и тако повећава брзину транспирације. |
| Ветар | У мирном ваздуху, вода изгубљена услед транспирације може се акумулисати у облику паре близу површине листа. Ово смањује стопу губитка воде, пошто је градијент потенцијала воде од унутрашњег ка спољашњем делу листа тада нешто мањи. Ветар одува велики део ове водене паре близу површине листа, чинећи потенцијални градијент стрмијим и убрзавајући дифузију молекула воде у околни ваздух. Чак и на ветру, међутим, може доћи до акумулације водене паре у танком граничном слоју ваздуха који се спорије креће поред површине листа. Што је ветар јачи, то ће овај слој тежити да буде тањи, а градијент потенцијала воде стрмији. |
| Снабдевање водом | Недостатак воде узрокован ограниченим снабдевањем водом из тла може довести до затварања стомата и смањења стопе транспирације. |
-
 Утицај температуре на брзину транспирације биљака.
Утицај температуре на брзину транспирације биљака. -
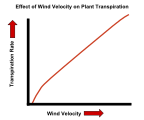 Утицај брзине ветра на брзину транспирације биљака.
Утицај брзине ветра на брзину транспирације биљака. -
 Утицај влаге на брзину транспирације биљака.
Утицај влаге на брзину транспирације биљака.




Током сезоне раста, лист транспирише много пута више воде од сопствене тежине. Јутар кукуруза испушта око 3.000–4.000 галона (11.400–15.100 литара) воде сваког дана, а велики храст може да испусти 40.000 галона (151.000 литара) годишње. Стопа транспирације је однос масе транспириране воде према маси произведене суве материје; коефицијент транспирације усева има тенденцију да падне између 200 и 1000 (i.e. биљке усева транспирују 200 до 1000 kg воде за сваки kg произведене суве материје).[10]
Стопе транспирације биљака могу се мерити бројним техникама, укључујући потометере, лизиметре, порометре, системе фотосинтезе и термометричке сензоре протока сока. Мерења изотопа показују да је транспирација већа компонента евапотранспирације.[11] Недавни докази из једне глобалне студије[12] стабилних изотопа у води показују да се транспирисана вода изотопски разликује од подземних вода и потока. Ово сугерише да вода у тлу није тако добро измешана као што се широко претпоставља.[13]
Кавитација[уреди | уреди извор]
Да би се одржао градијент притиска неопходан да би биљке остале здраве, оне морају континуирано да уноси воду својим коренима. Оне морају да буду у стању да задовоље потребе воде изгубљене због транспирације. Ако биљка није у стању да унесе довољно воде да остане у равнотежи са транспирацијом, долази до појаве познате као кавитација.[14] Кавитација се јавља када биљка не може да снабде свој ксилем адекватном количином воде, те уместо да се испуни водом, ксилем почиње да се пуни воденом паром. Ове честице водене паре се спајају и формирају блокаде унутар ксилема биљке. Ово спречава биљку да транспортује воду кроз свој васкуларни систем.[15] Не постоји очигледан образац где се кавитација јавља у ксилему биљке. Ако се тај проблем ефикасно не разреши, кавитација може довести до тога да биљка достигне своју трајну тачку увенућа и умре. Према томе, биљка мора да има метод којим ће уклонити ову блокаду кавитације, или мора да створи нову везу васкуларног ткива у целој биљци.[16] Биљка то чини затварањем својих стомата преко ноћи, што зауставља ток транспирације. Ово онда омогућава коренима да генеришу притисак од преко 0,05 mPa, а то је у стању да уништи блокаду и поново напуни ксилем водом, поново повезујући васкуларни систем. Ако биљка није у стању да створи довољан притисак да искорени блокаду, она мора спречити ширење блокаде употребом крушке коштице, а затим створити нови ксилем који може поново да повеже васкуларни систем биљке.[17]
Референце[уреди | уреди извор]
- ^ Cummins, Benjamin (2007), Biological Science (3 изд.), Freeman, Scott, стр. 215
- ^ а б Taiz, Lincoln (2015). Plant Physiology and Development. Sunderland, MA: Sinauer Associates, Inc. стр. 101. ISBN 978-1-60535-255-8.
- ^ Sinha, Rajiv Kumar (2004-01-01). Modern Plant Physiology (на језику: енглески). CRC Press. ISBN 978-0-8493-1714-9.
- ^ Benjamin Cummins (2007), Biological Science (3 изд.), Freeman, Scott, стр. 215
- ^ Freeman, Scott (2014). Biological Sciences. United States of America: Pearson. стр. 765—766. ISBN 978-0-321-74367-1.
- ^ Simon, E.J., Dickey, J.L, & Reece, J.B. (2019). Campbell essential biology. 7th New York: Pearson
- ^ Graham, Linda E. (2006). Plant Biology. Upper Saddle River, NJ 07458: Pearson Education, Inc. стр. 200–202. ISBN 978-0-13-146906-8.
- ^ „Capillary Action – Liquid, Water, Force, and Surface – JRank Articles”. Science.jrank.org. Архивирано из оригинала 2013-05-27. г. Приступљено 2013-06-18.
- ^ Mellander, Per-Erik; Bishop, Kevin; Lundmark, Tomas (2004-06-28). „The influence of soil temperature on transpiration: a plot scale manipulation in a young Scots pine stand”. Forest Ecology and Management (на језику: енглески). 195 (1): 15—28. ISSN 0378-1127. doi:10.1016/j.foreco.2004.02.051.
- ^ Martin, J.; Leonard, W.; Stamp, D. (1976), Principles of Field Crop Production (3rd изд.), New York: Macmillan Publishing Co., ISBN 978-0-02-376720-3
- ^ Jasechko, Scott; Sharp, Zachary D.; Gibson, John J.; Birks, S. Jean; Yi, Yi; Fawcett, Peter J. (3. 4. 2013). „Terrestrial water fluxes dominated by transpiration”. Nature. 496 (7445): 347—50. Bibcode:2013Natur.496..347J. PMID 23552893. S2CID 4371468. doi:10.1038/nature11983.
- ^ Evaristo, Jaivime; Jasechko, Scott; McDonnell, Jeffrey J. (2015-09-03). „Global separation of plant transpiration from groundwater and streamflow”. Nature. 525 (7567): 91—94. Bibcode:2015Natur.525...91E. ISSN 0028-0836. PMID 26333467. S2CID 4467297. doi:10.1038/nature14983.
- ^ Bowen, Gabriel (2015-09-03). „Hydrology: The diversified economics of soil water”. Nature. 525 (7567): 43—44. Bibcode:2015Natur.525...43B. ISSN 0028-0836. PMID 26333464. S2CID 205086035. doi:10.1038/525043a.
- ^ Zhang, Yong-Jiang (децембар 2016). „Reversible Leaf Xylem Collapse: A Potential "Circuit Breaker" against Cavitation”. Plant Physiology. 172 (4): 2261—2274. PMC 5129713
 . PMID 27733514. doi:10.1104/pp.16.01191.
. PMID 27733514. doi:10.1104/pp.16.01191.
- ^ Hochberg, Uri (јун 2017). „Stomatal Closure, Basal Leaf Embolism, and Shedding Protect the Hydraulic Integrity of Grape Stems”. Plant Physiology. 174 (2): 764—775. PMC 5462014 . PMID 28351909. doi:10.1104/pp.16.01816.
- ^ Holbrook, Michele (мај 2001). „In Vivo Observation of Cavitation and Embolism Repair Using Magnetic Resonance Imaging”. Plant Physiology. 126 (1): 27—31. PMC 1540104 . PMID 11351066. doi:10.1104/pp.126.1.27.
- ^ Tiaz, Lincoln (2015). Plant Physiology and Development. Massachusetts: Sinauer Associates, Inc. стр. 63. ISBN 978-1605352558.
Спољашње везе[уреди | уреди извор]
- Transpiration by Trees
- USGS The Water Cycle: Evapotranspiration Архивирано на сајту Wayback Machine (7. децембар 2013)
| Државне | |
|---|---|
| Остале | |