
Rđa (biologija)
| Rđe | |
|---|---|
.jpg)
| |
| Puccinia recondita na pšeničnom listu | |
| Naučna klasifikacija | |
| Domen: | Eukaryota |
| Carstvo: | Fungi |
| Tip: | Basidiomycota |
| Klasa: | Pucciniomycetes |
| Red: | Pucciniales |
| Porodice | |
Rđe su biljne bolesti uzrokovane patogenim gljivama reda Puciniales (ranije poznate kao Uredinales).
Procenjuje se da je trenutno prihvaćeno 168 rodova rđe i približno 7000 vrsta, od kojih više od polovine pripada rodu Puccinia [1]. Gljive rđe su visoko specijalizovani biljni patogeni sa nekoliko jedinstvenih karakteristika. Grupno uzete, gljive rđe su raznolike i pogađaju mnoge vrste biljaka. Međutim, svaka vrsta ima vrlo uski raspon domaćina i ne može se preneti na biljke koje nisu domaćini. Pored toga, većina gljivica rđe ne može se lako uzgajati u čistoj kulturi.
Jedna vrsta gljiva rđe može zaraziti dva različita biljna domaćina u različitim fazama svog životnog ciklusa i može proizvesti do pet morfološki i citološki različitih struktura koje proizvode spore, odnosno spermogoniju, aeciju, urediniju, teliju i bazidiju u uzastopnim fazama razmnožavanja[2]. Svaka spora je vrlo specifična za domaćina i obično može zaraziti samo jednu vrstu biljaka.
Gljive rđe su obvezni biljni patogeni koji zaraze samo žive biljke. Infekcije počinju kada spora sleti na površinu biljke, klija i napada svog domaćina. Infekcija je ograničena na biljne delove kao što su lišće, peteljke, nežni izdanci, stabljika, plodovi itd[1]. Biljke sa ozbiljnom infekcijom rđe mogu izgledati kržljavo, hlorotično (požutelo) ili mogu pokazivati znakove infekcije poput plodnih tela rđe. Gljive rđe rastu unutarćelijski i stvaraju plodišta koja proizvode spore unutar ili, češće, na površinama pogođenih biljnih delova. Neke vrste rđe formiraju višegodišnje sistemske infekcije koje mogu prouzrokovati deformacije biljaka kao što su zaostajanje u rastu, veštičina metla, rak stabljike, galice ili hipertrofija pogođenih biljnih delova.
Rđe su ime dobile jer ih se najčešće primećuje kao naslage praškastih rđastih ili smeđih spora na biljnim površinama. Rimski poljoprivredni festival Robigalia (25. aprila) drevno je poreklo u borbi protiv pšenične rđe[3].
Uticaji[uredi | uredi izvor]
Rđe se smatraju najštetnijim patogenima za poljoprivredu, hortikulturu i šumarstvo. Gljive rđe su glavna briga i ograničavajući faktori uspešnog uzgajanja poljoprivrednih i šumskih kultura. Rđa belog bora, rđa pšeničnih stabljika, rđa soje i rđa kafe primeri su velike štetne pretnje važnim usevima sa ekonomskog stanovišta[1].
Životni ciklus[uredi | uredi izvor]
Sve rđe su obvezni paraziti, što znači da im je potreban živi domaćin da završi svoj životni ciklus. Rđe uglavnom ne ubijaju biljku domaćina, ali mogu ozbiljno smanjiti rast i prinos[4]. Usevi žitarica mogu biti uništeni u jednoj sezoni; stabla hrasta zaražena u prvih pet godina rđom Cronartium quercuum često umiru[5].
_-Osterloh-_-Brendel_10_h,_2-.jpg)
Gljive rđe mogu da proizvedu do pet vrsta spora od odgovarajućih tipova plodišta tokom svog životnog ciklusa, u zavisnosti od vrste.
Morfološki tipovi gljiva rđe
- 0 - Picniospores ( Spermatia ) iz Picnidia. Oni služe uglavnom kao haploidne polne ćelije u heterotalnim rđama .
- I - Aeciospore iz Aecije. One uglavnom služe kao neponavljajuće dikariontske, aseksualne spore i nastavljaju da zaraze primarnog domaćina.
- II - Urediniospore iz Uredia ( Uredinia ). Oni služe kao ponavljajuće dikariontske vegetativne spore. Ove spore se nazivaju fazom koja se ponavlja, jer mogu da izazovu auto-infekciju na primarnom domaćinu, ponovo zarazeći istog domaćina na kome su nastale spore. Često su obilne, crvene / narandžaste i istaknuti znak bolesti rđe.
- III - Teliospore iz Telije . Ove dikariotske spore su često faza preživljavanja / prezimljavanja životnog ciklusa. Obično ne zaraze biljku direktno; umesto toga klijaju da bi proizveli bazidiju i bazidiospore.
- IV - Basidiospore iz Teliospore. Ove haploidne spore vetrom često zaraze alternativnog domaćina u proleće.[6][7] Retko se primećuju van laboratorije .
Gljive rđe često su kategorisane prema životnom ciklusu. Tri osnovna tipa životnih ciklusa prepoznaju se na osnovu broja vrsta spora kao makrociklični, demiciklični i mikrociklični[1].
Makrociklični životni ciklus ima sva spora, demicikličkom nedostaje uredinijalno stanje, a mikrocikličnom ciklusu osnovno, piknijalno i ecijalno stanje, pa poseduju samo urediniju i teliju. Spermagonija može biti odsutna kod svake vrste, ali posebno u mikrocikličnom životnom ciklusu. U makrocikličnim i demicikličnim životnim ciklusima, rđa može biti ili domaćin naizmeničan (heteroecious) (tj. Aecijsko stanje je na jednoj vrsti biljke, a telialno stanje na drugoj i nepovezanoj biljci), ili naizmenično ( autoecious ) (bez domaćina) ( tj. vazdušna i telijalna stanja na istom biljnom domaćinu).
Heteroetične gljive rđe zahtevaju da dva nepovezana domaćina završe svoj životni ciklus, pri čemu su primarni domaćini zaraženi aeciosporama, a alternativni domaćin zaraženi bazidiosporama. To se može suprotstaviti auto-domaćoj gljivi koja može završiti sve delove svog životnog ciklusa na jednoj vrsti domaćina[6]. Razumevanje životnih ciklusa gljivica rđe omogućava pravilno upravljanje bolestima[8].
Odnos biljke domaćina i gljivice[uredi | uredi izvor]
Postoje određeni obrasci odnosa sa biljnim grupama domaćinima i gljivicama rđe koje na njima parazitiraju. Neki rodovi gljivica rđe, naročito Pucinia i Uromices, sadrže vrste koje su sposobne da parazitiraju na biljkama mnogih porodica. Čini se da su ostali rodovi rđe ograničeni na određene biljne grupe. Ograničenje domaćina može se, kod heteroetičnih vrsta, primeniti na obe faze životnog ciklusa ili samo na jednu fazu[1].
Proces zaraze[uredi | uredi izvor]
Spore gljivica rđe mogu se raširiti vetrom, vodom ili insektima[9]. Kada spora naiđe na osetljivu biljku, ona može klijati i zaraziti biljna tkiva. Spore rđe obično klijaju na biljnoj površini, izrastajući kratku hifu koja se naziva zametna cev . Ova klica cev može locirati Stoma od strane odgovornog procesa dodir poznat kao higmotropim. To uključuje orijentaciju na grebene stvorene od ćelija epiderme na površini lista i raste usmereno dok ne naiđe na stomu[10].

Preko stome, hifalni vrh stvara infektivnu strukturu koja se naziva apresorijum . Sa donje strane apresorija, tanka hifa raste nadole da zarazi biljne ćelije[11]. Smatra se da je čitav proces posredovan osetljivim na istezanje kanalom kalcijumovih jonskih kanala smeštenih na vrhu hife, koji proizvode električne struje i menjaju gen izraz, podstičući formiranje apresorijuma[12].
Jednom kada je gljiva napala biljku, ona prerasta u biljne mezofilne ćelije, proizvodeći specijalizovane hife poznate kao haustorija. Haustorija prodire u ćelijske zidove, ali ne i kroz ćelijske membrane: biljne ćelijske membrane invaginiraju oko glavnog haustorijskog tela formirajući prostor poznat kao ekstra haustorska matrica. Vrpca bogata gvožđem i fosforom premošćuje biljne i gljivične membrane u prostoru između ćelija za protok vode, poznatom kao apoplast, sprečavajući tako hranljive materije da dospeju do ćelija biljke. Haustorium sadrži aminokiselinske i Heksoza šećera transportere i H+ - ATPases koje se koriste za aktivnim transportom hranljivih materija iz postrojenja, hrani gljivice[13]. Gljiva nastavlja da raste, prodirući u sve više biljnih ćelija, sve dok se ne dogodi rast spora . Proces se ponavlja svakih 10 - 14 dana, proizvodeći brojne spore koje se mogu proširiti na druge delove iste biljke ili na nove domaćine.
Uobičajene gljive rđe u poljoprivredi[6][8][14][uredi | uredi izvor]
- Kronartium ribicola ( rđa belog bora) ; primarni domaćini su ribizla, a beli borovi sekundarni. Heterociklični i makrociklični
- Gimnosporangium juniperi-virginianae (rđa od kedra i jabuka) ; Juniperus virginiana je primarni (telijalni) domaćin, a jabuka, kruška ili glog sekundarni ( vazdušni ) domaćin. Heteroecious i demicikličan
- Hemileia vastatrik (rđa od kafe) ; Primarni domaćin je biljka kafe; nepoznati alternativni domaćin. Heteroecious
- Phakopsora meibomiae i P. pachirhizi ( sojina rđa ) ; Primarni domaćin je soja i razne mahunarke. Nepoznati alternativni domaćin. Heteroecious
- Pucinia coronata (krunska rđa ovsa i ljuske) ; Zob je primarni domaćin; Rhamnus spp. (Buckthorn) je alternativni domaćin. Heteroecious i macrociclic
- Pucinia graminis ( matična rđa pšenice i Kentucki bluegrass, ili crna rđa žitarica ); Primarni domaćini su: plava trava iz Kentuckija, ječam i pšenica; Obična žutika je alternativni domaćin. Heteroecious i macrociclic
- Pucinia hemerocallidis (rđa Dailili ) ; Dailili je primarni domaćin; Patrina sp je alternativni domaćin. Heteroecious i macrociclic
- Pucinia triticina (rđa smeđe pšenice) u zrnu
- Pucinia sorghi (obična rđa kukuruza) [15]
- Pucinia striformis (žuta rđa) žitarica
- Uromices apendiculatus u pasulju (Phaseolus vulgaris) [16]
- Pucinia melanocephala (smeđa rđa šećerne trske)
- Pucinia kuehnii (narandžasta rđa šećerne trske )
Upravljanje bolestima gljivica rđe[uredi | uredi izvor]
Metode suzbijanja bolesti gljivica rđe u velikoj meri zavise od životnog ciklusa određenog patogena. Slede primeri primene planova za upravljanje bolestima koji se koriste za kontrolu makrocikličnih i demicikličnih bolesti:
Makrociklična bolest: Izrada plana upravljanja za ovu vrstu bolesti u velikoj meri zavisi od toga da li se faza ponavljanja (urediniospore) javlja na ekonomski važnoj biljci domaćinu ili na alternativnom domaćinu. Na primer, faza ponavljanja bolesti rđe od mehura od belog bora ne javlja se na belim borovima, već kod alternativnog domaćina, Ribizli. Tokom avgusta i septembra Ribizle daju teliospore koje inficiraju bele borove. Uklanjanje alternativnog domaćina narušava životni ciklus gljivica rđe Kronartium ribicola, sprečavajući stvaranje bazidiospora koje zaraze primarnog domaćina. Iako spore iz belih borova ne mogu da zaraze druge bele borove, spore za preživljavanje mogu prezimiti na zaraženim borovima i ponovo zaraziti Ribizle. sledeće sezone. Zaraženo tkivo uklanja se iz belih borova i strogih karantina Ribizli održavaju se u oblastima visokog rizika[6][17].
Pucinia graminis je makrociklična heteroecious gljiva koja uzrokuje bolest rđe na stabljici pšenice. Faza ponavljanja ove gljive javlja se na pšenici, a ne na alternativnom domaćinu, žutiku . Faza ponavljanja omogućava da bolest opstane u pšenici, iako se alternativni domaćin može ukloniti. Sadnja otpornih useva idealan je oblik prevencije bolesti, međutim, mutacije mogu dovesti do novih sojeva gljivica koji mogu savladati otpornost biljaka. Iako se bolest ne može zaustaviti uklanjanjem alternativnog domaćina, životni ciklus je poremećen i stopa mutacije je smanjena zbog smanjene genetske rekombinacije. To omogućava usevima uzgojenim otporom da ostanu efikasni tokom dužeg vremenskog perioda[6][18].
Demiciklična bolest: Budući da se u životnom ciklusu demicikličnih gljiva ne ponavlja faza, uklanjanje primarnog ili alternativnog domaćina poremetiće ciklus bolesti. Ova metoda, međutim, nije visoko efikasna u lečenju svih demicikličnih bolesti. Na primer, bolest rđe od kedrovine i jabuke može potrajati uprkos uklanjanju jednog od domaćina, jer se spore mogu širiti sa velike udaljenosti. Ozbiljnošću bolesti rđe kedra i jabuke može se upravljati uklanjanjem bazidiospore koje proizvode gale iz smreke ili primenom zaštitnih fungicida na smreke[19].
Kućna kontrola[uredi | uredi izvor]
Bolesti rđe su veoma teške za lečenje. Fungicidi kao što su Mancozeb ili Triforine može da pomogne, ali nikada ne može da iskoreni bolest. Dostupna su neka organska preventivna rešenja, a poznato je da sumporni prah zaustavlja klijanje spora. Visoki higijenski standardi, dobra drenaža tla i pažljivo zalivanje mogu smanjiti probleme. Bilo koja pojava rđe mora se odmah rešiti uklanjanjem i spaljivanjem svih pogođenih listova. Kompostiranje ili ostavljanje zaražene vegetacije na zemlji širiće bolest.
Komercijalna kontrola[uredi | uredi izvor]
U nekim velikim usevima fungicidi se primenjuju vazduhom. Proces je skup i primena fungicida je najbolje rezervisana za sezone kada su folijarne bolesti ozbiljne. Istraživanja pokazuju da što je veća ozbiljnost folijarne bolesti, to je veći povraćaj od upotrebe fungicida[20]. Južna bolest kukuruzne rđe, može se zameniti sa uobičajenom rđom. Karakteristična karakteristika južne rđe je ta što se pustule uglavnom formiraju na gornjoj površini lista, a spore su više narandžaste boje. Južna rđa se brže širi i ima veći ekonomski uticaj kada i dalje postoje topli i vlažni vremenski uslovi. Pravovremene primene fungicida za suzbijanje južne rđe su presudnije nego kod obične rđe[21],
Za bolesti rđe mogu se primeniti razne preventivne metode:
- Visok nivo vlage može pogoršati simptome bolesti rđe. Izbegavanje nadzemnog zalivanja noću, navodnjavanjem kap po kap, smanjenjem gustine useva i upotrebom ventilatora za cirkulaciju vazdušnog toka može smanjiti težinu bolesti.
- Upotreba biljnih sorti otpornih na rđu
- Plodored useva može prekinuti ciklus bolesti jer su mnoge rđe specifične za domaćina i ne održavaju se dugo bez svog domaćina.
- Inspekcija uvoznih biljaka i sečenja na simptome. Važno je kontinuirano posmatrati biljke, jer bolesti rđe imaju latentni period (biljka ima bolest, ali ne pokazuje simptome).
- Upotreba semena bez bolesti može smanjiti pojavu nekih rđa [18]
Pogođene biljke domaćini[uredi | uredi izvor]
Verovatno je da su neke vrste rđe pogođene većinom biljnih vrsta. Rđe su često nazvane po vrsti domaćinu koju zaraze. Na primer; Pucinia kanthii inficira cvetnu biljku koktel ( Ksanthium ). Nedavno je prijavljeno ukupno 95 gljivica rđe koje pripadaju 25 rodova povezanih sa 117 vrsta šumskih biljaka koje pripadaju 80 rodova domaćina pod 43 porodice domaćina sa zapadnih Gata, Kerala, Indija[1].
Gljive rđe uključuju[uredi | uredi izvor]
Rodovi zaraženi rđom uključuju[1][uredi | uredi izvor]
- Acacia
- Acalypha
- Ageratina
- Albizia
- Arundinaria
- Bambusa
- Bidens
- Blepharis
- Bombax
- Bridelia
- Callicarpa
- Canarium
- Canthium
- Catunaragam
- Cinnamomum
- Cissus
- Cleistanthus
- Clerodendron
- Coffea
- Coix
- Cosmostigma
- Crotalaria
- Dalbergia
- Dendrocalamus
- Derris
- Diospyros
- Dipterocanthus
- Elaeagnus
- Elephantopus
- Elettaria
- Eragrostis
- Euphorbia
- Ficus
- Flacourtia
- Grewia
- Holarrhena
- Holoptelia
- Hypericum
- Ichnocarpus
- Ischaemum
- Jasminum
- Justicia
- Loesneriella
- Luvunga
- Meiogyne
- Meliosma
- Mimusops
- Morus
- Neolitzea
- Ocimum
- Olea
- Oxalis
- Pavetta
- Persicaria
- Phyllanthus
- Plectranthus
- Plumeria
- Pongamia
- Premna
- Protasparagus
- Rubus
- Salix
- Spondia
- Strobilanthes
- Strychnos
- Tabernaemontana
- Terminalia
- Toddalia
- Trichosanthes
- Vernonia
- Vigna
- Wrightia
- Xanthophyllum
- Xylia
- Ziziphus
Neki od poznatijih domaćina uključuju[uredi | uredi izvor]
- Arisaema triphyllum, Jack-in-the-pulpit
- Avena sativa, Oats
- Berberis vulgaris, Šimširika
- Vicia faba - Bob (biljka)
- Coffea arabica - Kafa
- Crataegus monogyna - Hawthorn
- Chrysanthemum
- Cydonia - Quince
- Euphorbia maculata, Spotted Spurge
- Fuchsia spp,
- Garlic - Beli luk
- Hordeum vulgare, Barley
- Juniperus virginiana, Red Cedar (Juniper apple disease)
- Juniperus communis - Kleka (vrsta)
- Allium ampeloprasum - Leek
- Malus – Jabuka
- Mentha piperita - Peppermint
- Mespilus - Mušmula
- Onion - Crni luk
- Pelargonium
- Primula veris
- Primula vulgaris
- Pyrus - Pear
- Rosa spp, Ruža
- Triticum spp., Wheat
- Oxalis spp., Oxalis
- Secale cereale, Rye
- Senecio vulgaris - Common groundsel
- Xanthium canadense Cocklebur
Hiperparaziti rđe[uredi | uredi izvor]
U porodici Sfaeropsidaceae gljiva Sfaeropsidales, vrste roda Darluca su hiperparaziti na rđama.[22]
Galerija[uredi | uredi izvor]
-
 Gljiva rđe na listu, pod malim uvećanjem.
Gljiva rđe na listu, pod malim uvećanjem. -
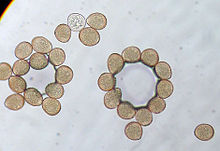 Urediniospore gljivice rđe.
Urediniospore gljivice rđe. -
 Dijagram koji predstavlja proces zaraze gljivicama rđe
Dijagram koji predstavlja proces zaraze gljivicama rđe -
 Rđa na luku
Rđa na luku
_Pengo.jpg)
_pustules_of_urediniospores_Pengo.jpg)


Vidi još[uredi | uredi izvor]
- Matična rđa
- Rđa listova pšenice
- Lisna rđa (ječam)
- Gljivice
- Smut (gljivica)
- Rđa soje
- Rust (programski jezik) (nazvan po gljivici Rust)
Reference[uredi | uredi izvor]
- ^ a b v g d đ e Mohanan C. (2010). Rust Fungi of Kerala. Kerala, India: Kerala Forest Research Institute. str. 148. ISBN 978-81-85041-72-8.
- ^ Kolmer, James A; Ordonez, Maria E; Groth, James V (2001). eLS (na jeziku: engleski). John Wiley & Sons, Ltd. ISBN 9780470015902. S2CID 1434349. doi:10.1002/9780470015902.a0021264.
- ^ Evans, R. (2007). Utopia Antiqua: Readings of the Golden Age and Decline at Rome. Taylor & Francis. ISBN 978-1-134-48787-5. Pristupljeno 2018-01-12.
- ^ Central Science Laboratory. (2006). Plant Healthcare: Rusts [Fact Sheet]. Retrieved from www.csldiagnostics.co.uk
- ^ „Rust Fungi”. www.backyardnature.net.
- ^ a b v g d Schumann, G. & D'Arcy, C. (2010). Essential plant pathology. APS Press
- ^ Scott, K.J, & Chakravorty, A.K., (1982), The Rust fungi. Academic Press.
- ^ a b Petersen, Ronald H. (1974). „The Rust Fungus Life Cycle”. The Botanical Review. 40 (4): 453—513. S2CID 45598137. doi:10.1007/BF02860021..
- ^ Craigie, J.H. (1931). Phytopathology, 21,1001
- ^ Dickinson, M. Molecular Plant Pathology. 2003.
- ^ Deising, H.B., S. Werner, and M. Wernitz, The role of fungal appressoria in plant infection. Microbes Infect, 2000. 2(13) p. 1631-41.
- ^ Zhou, X.L., et al., A mechanosensitive channel in whole cells and in membrane patches of the fungus Uromyces. Science, 1991. 253(5026) p. 1415.
- ^ Voegele, R.T. and K. Mendgen, Rust haustoria: nutrient uptake and beyond. New Phytologist, 2003. 159(1) p. 93-100.
- ^ Cornell University. (2010). Daylily rust: Puccinia hemerocallidis [Fact sheet]. Retrieved from http://plantclinic.cornell.edu
- ^ Hooker, Arthur L (1967). „The Genetics and Expression of Resistance in Plants to Rusts of the Genus Puccinia”. Annu. Rev. Phytopathol. 5 (1): 163—178. doi:10.1146/annurev.py.05.090167.001115.
- ^ Hurtado-Gonzales, O. P.; Valentini, G.; Gilio, T. A.; Martins, A. M.; Song, Q.; Pastor-Corrales, M. A. (2016). „Fine Mapping of Ur-3, a Historically Important Rust Resistance Locus in Common Bean”. G3 (Bethesda, Md.). 7 (2): 557—569. PMC 5295601
 . PMID 28031244. doi:10.1534/g3.116.036061.
. PMID 28031244. doi:10.1534/g3.116.036061.
- ^ Cornell University. (2005). White Pine Blister Rust: Cronartium ribicola [Fact sheet]. Retrieved from http://plantclinic.cornell.edu
- ^ a b Marsalis, M. & Goldberg, N. (2006). Leaf, Stem, And Stripe Rust Diseases of Wheat. [Fact sheet]. New Mexico State University
- ^ Wallis, C. & Lewandowski, D. (2008). Cedar Rust Diseases of Ornamental Plants. [Fact Sheet]. Ohio State University
- ^ „Stopsoybeanrust.com”. www.stopsoybeanrust.com. Arhivirano iz originala 12. 06. 2018. g. Pristupljeno 06. 09. 2020.
- ^ „Common Corn Rust”. www.channel.com.
- ^ faculty.ucr.edu (retrieved December 2015)
Literatura[uredi | uredi izvor]
- Mohanan C. (2010). Rust Fungi of Kerala. Kerala, India: Kerala Forest Research Institute. str. 148. ISBN 978-81-85041-72-8.
Spoljašnje veze[uredi | uredi izvor]
| Državne | |
|---|---|
| Ostale | |